
The Pholadidae are a family of marine bivalves adapted for boring. They have neither hinge-teeth nor external ligament, and are characterised by having externally placed anterior adductor muscles, and accessory plates (plaxes) in addition to the normal bivalve shell; for this reason, some early authors classed them with the Multivalves. The family is almost entirely marine, and is closely related to the ship-worms (Family Teredinidae). Like them, they are without hinge-teeth but have a blade-like apophysis, for the attachment of the pedal muscles, projecting inwards from the umbo.
Few Pholads are of economic importance, though Martesia striata, a wood-borer, is more destructive than Teredo in the Philippines, and Xylophaga, another wood-boring genus, occasionally attacks submarine cables. Diplothyra smithii and Penitella conradi are both shell-borers, in oysters and Haliotis respectively, but neither appears to do any harm to the host.
A few species have occasionally been used for food, but none is of commercial importance as most are too difficult to remove from their burrows, and arc too full of sand to be palatable. Pholas dactylus was used for food by the Romans, and the large Oyrtopleura costata was formerly sold in Cuban markets, one specimen making a meal for two people.
Diagrams to show the terms used in describing a Pholad shell
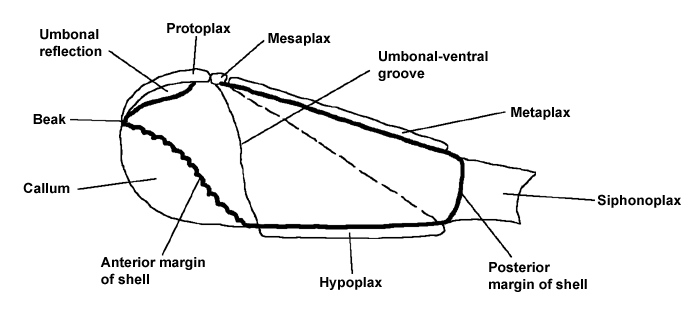
External view of left valve to show the arrangement of the accessory plates
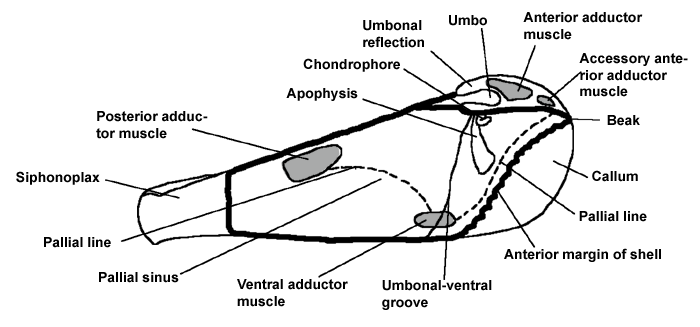
Internal view of left valve to show the arrangement of the muscle scars etc
The family is world-wide in distribution, attaining its maximum development in the East Pacific. The vertical range is mainly less than 250 fm., though a few abyssal species have been recorded. Others are pelagic, being found in floating wood. The genera are locally restricted by their specialised habitat requirements. Martesia and Xylophaga are generally limited to wood, Zirfaea prefers salt marsh peat and stiff mud, Barnea and Fholadidea bore into clays and sandstones, while Pholas may attack gneiss. Stunted specimens (stenomorphs) are produced in overcrowded conditions and in very hard substrates.
At one time there was considerable speculation about the mechanics of the boring process. Some malacologists believed that boring was accomplish¬ed by an acid secretion, others that the foot did the boring using embedded siliceous spicules, while many considered that the shell was the tool. The belief that an acid is employed is no longer held, and it is now recognised that the burrow is produced by the shell manipulated by the specially modified adductor muscles and aided by the foot, siphons, mantle, water and accessory sand grains, all of which contribute to the process of abrasion. The shells are of aragonite and mechanically are capable only of boring substrata softer than themselves, though the harder rocks may be bored where they are friable or contain softer elements.
In some genera (e.g. Pholas, Xylophaga) there is no basic change in shell morphology from juvenile to adult; in others, however, (e.g. Martesia, Pholadidea) there are two distinct stages of shell growth. The young shell is beaked and widely gaping anteriorly, while in the adult the anterior gape is closed by a calcareous callum. The young stage (working-, or Zirfaea-stage) continues as long as the animal is actively boring.
When excavation is completed, the foot atrophies, the pedal gape is closed by the callum and the various plaxes develop. The resting-stage has then been attained. In the callum-forming Pholads, the animal stores a considerable amount of calcium, enabling the callum to be laid down very rapidly. Thus very few individuals are found with a partial callum, and consequently the young stages have occasionally been described as separate genera. Since the adults with complete callum vary considerably in size, depending on the hardness of the substrate, it was formerly thought that resorption could occur, followed by further active growth with eventual re¬formation of the callum. This is not so, for the foot atrophies when the callum is produced and can no longer contribute to the enlargement of the burrow. The muscles used in boring do not atrophy as a slight movement of the valves is maintained to permit a circulation of water within the burrow.
| 1 | a) Adult shell with callum | 2 |
| b) Adult shell without callum | 3 | |
| 2 | a) With chitinous accessory plates around siphons; usually boring in sandstone or clay; S.W. England and Ireland | Pholadidea loscombiana |
| b) With funnel-shaped pits below umbonal reflexion; usually restricted to drift-wood; rare, in S.W. England | Martesia striata | |
| 3 | a) Shell almost globular, without apophyses; boring in wood | 7 |
| b) Shell elongated; boring in mud, clay or rock | 4 | |
| 4 | a) Shell with umbonal-ventral groove; on all British coasts, but rather local | Zirfaea crispata |
| b) Shell without umbonal-ventral groove | 5 | |
| 5 | a) Umbonal reflexion septate; shell up to 5" or more in length; S. and W, coasts | Pholas dactylus |
| b) reflexion simple; shell generally less than 3" long | 6 | |
| 6 | a) Anterior end beaked and gaping | Barnea parva |
| b) Anterior end rounded | Barnea Candida | |
| 7 | a) Shell up to 1" in length, with conspicuous furrow from umbo to anterior angle; rare | Xylophaera craestans |
| b) Shell under without furrow | Xylophaga dorsalis |
Acknowledgement; The author. Dr. J. Bowden of Glasgow University, and the publishers, Oliver and Boyd Ltd., are thanked for permission to use the key, based on one due to appear in 'British Bivalvia’.