
Introduction
The Farasan Islands are located in the Red Sea about 40 km off the Arabian coast, opposite the town of Jizan in the extreme southwest of Saudi Arabia. This archipelago is situated on the remarkably shallow Farasan Bank and comprises more than 300 islands, islets and shoals of which only three (Farasan Kebir (400 km²), Qummah (15 km²) and As Saqid (160 km²) (figure 1)) are permanently inhabited. Geologically, the Farasan Islands consist of an originally more or less uniform flat fossil coral reef that rose 0–30 m above sea level during the late Pliocene to early Pleistocene (McFayden, 1930).

figure 1: Aerial image showing the sampling location at Abu Tok on As Saqid Island, Farasan Archipelago. A shallow bank, consisting of algae beds and reefs, connects As Saqid to Safrah Island. In the background Khur Abu Tok, Saghir ar-Ryaq and Kabir ar-Riyaq Islands which connect to Safrah Island by a coral reef.
Tectonic processes led to shattering and uplift. The water around the islands is less than 100 m deep and various marine habitats are represented including sea grass beds, mangroves and vast areas of fringing reefs. The high salinity of the water surrounding the Farasan Islands is a consequence both of the hot climate and the absence of any freshwater input. The surface temperature ranges, according to season, from approximately 25˚ to 31˚C. Temperature declines with increasing depth to about 21.5˚C at 700 m where it remains remarkably constant. The normal tides are small, with peak tides of about 0.5 m, but there are seasonal and geographical long-term variations in water level over longer periods (Bemert and Ormond, 1981). The seasonal tides, and the pattern of prevailing winds and currents, are influenced by monsoons in the Indian Ocean. During the winter the prevailing winds are from the south, and are responsible for the numerous molluscs found on southerly oriented beaches, especially on Farasan Kebir.
The marine and coastal environments are generally in a healthy condition and the level of human exploitation is low. There are, however, localized impacts arising from unsustainable fishing practices, tourism development and oil pollution. Water quality is expected to decline and water use is expected to increase rapidly in the near future. The region is regarded as having a high conservation value because of the diversity of marine habitats and their importance for marine mammals, turtles and seabirds, and the small artisanal fishery. In view of this, it was decided in 1988 to establish a marine protected area around the Farasan Islands (Gladstone, 1994, 2000, 2000a).
This region of the Red Sea is poorly documented and the marine malacofauna has not been studied. This article presents a preliminary species list of bivalve molluscs inhabiting the coastal waters of the Farasan Islands. Previous records of marine molluscs from Saudi Arabia’s Red Sea coast were published by Sharabati (1981). Dekker and de Ceuninck van Capelle (1994) carried out a survey of Yemen Red Sea shells collected during the Tibia-1 expedition in 1993. The most northern sampling locality (Zahrat Ashiq island) of their study area was about 50 km southeast of the area sampled in this study. Dekker and Orlin (2000) provide a checklist of all known marine molluscs of the Red Sea and adjacent areas. The findings of this study are compared to those of Dekker and de Ceuninck van Capelle (1994), Dekker and Orlin (2000) and the OBIS Indo-Pacific Molluscan Database (Rosenberg et al., 2004).

figure 2: A mangrove stand at Kharij as Sailah, growing in the deep crevices cutting into the coral rock of which the entire Farasan Archipelago consists.
Methods
Bivalve sampling was carried out from 27 March to 18 October 2009, at 18 localities on Farasan Kebir, As Saqid and Qummah islands. The shore in the study area is comprised predominantly of coral fringe reefs, lagoons and long sandy beaches. Occasionally there are small bays with Avicennia spp. or Rhizophora spp. mangroves (Kharij as Sailah, Khor Kandrah, Khor Farasan) (figure 2) or macro-algal beds and reefs off the shore line (Qummah, As Saqid West (figure 5)). The location of the sampling sites, their habitat type and species richness are shown in figure 4.

figure 3: Atrina vexillum (length c.22cm) Qummah Island (Farasan Archipelago).
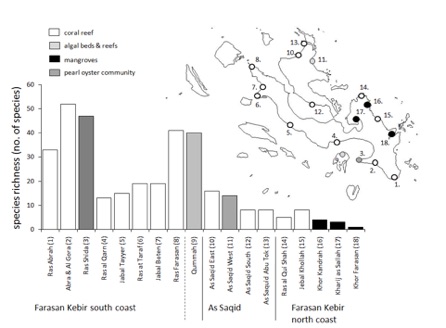
figure 4: The 18 sampling locations on Farasan Kebir, As Saqid and Qummah Islands, showing their habitat type and marine bivalve species richness.
Most shells were gathered by beach-combing (e.g. figure 3). When possible, the high and low water tide lines were inspected, and intermediate lines as far as they were marked by drift material. The sampling effort at most (10) sites was limited to 1 h; this was increased to 2 h at five sites (Ras Farasan, Ras al Qarn and Jabal Boten on Farasan Kebir; Qummah Island; and As Saqid West) and to 3 h at one site (Ras Abrah on Farasan Kebir). However, a disproportionate amount of time was spent at places with many shells and/or fine material. At Abrah and Al Gora Beach (south coast of Farasan Kebir) where shells were extraordinarily numerous due to the predominant wind direction and the open coast line, the sampling time was 31 h. At Ras Shida, where pearl divers leave their by-catch, the sampling time was 21 h.
Bivalves were also found by searching rock surfaces and by turning over stones embedded in sand or in rock pools. Occasionally, observations were made or samples were taken while snorkelling. Collecting methods did not include SCUBA-diving or dredging, and the search range was limited to the supra-littoral zone down to 2 m below the low water mark.
Species of boring or endo-symbiotic bivalves were collected from pieces of dead coral, sponges or submerged wood. Old and bleached shells that were brought from deeper waters by trawl, pearl and artisanal mollusc fisheries in former times were not included in the survey. Living specimens were determined on the spot and released afterwards; dead shell material was determined at the King Khalid Wildlife Research Centre (KKWRC), Thumamah, Saudi Arabia.
Bivalve species were determined using the following references: Sharabati (1984); Delsaerdt (1986); Oliver (1992); Dekker and Goud (1994); Bosch et al. (1995); Oliver and Chesney (1997); Vidal (1997); Oliver and Zuschin (2000); and Rusmore-Villaume (2008). The systematics and nomenclature used here predominantly follow Rosenberg et al. (2004).

figure 5: A shallow, sandy lagoon showing the sampling location at As Saqid West. Mangrove trees growing at the beach while farther outside algae beds and reefs fringe the lagoon.
Results and discussion
In total, 117 species of marine bivalves in 31 families were recorded during this study (table 1). Two species (Chlamys livida, Lima lima) are not listed by either Dekker and de Ceuninck van Capelle (1994) or Dekker and Orlin (2000) but are listed in Rosenberg et al. (2004).
According to Dekker and Orlin (2000), Chlamys livida is synonymous with Chlamys superficialis, but following Oliver (1992) the latter species has a microsculpture limited to the right valve and the anterior ear of the left valve lacks reticulate scaling. Since Rosenberg et al. (2004) separate these two species, and since in the samples collected during this study these characteristics could be distinguished, this article treats them as two species.
Two Lima species are listed in Dekker and Orlin (2000): Lima vulgaris and Lima paucicostata. The latter is considered a synonym of L. lima (Oliver, 1992). Since L. lima is the older name it is given priority. Other authors (e.g. Sowerby and Morris in Sharabati, 1984) distinguish L. paucicostata and L. lima on the basis of the number of ribs. The number of ribs varies between 11 and 18 in the material cited by Sharabati (1984) and until more material is examined it appears not justified to assign specific rank to L. paucicostata (Oliver, 1992).
Dekker and Orlin (2000) synonymise Spondylus groschi with Spondylus marisrubri but consider Spondylus spinosus a distinct species, being a senior synonym of S. zonalis and S. linguafelis. Contrary to that, Oliver (1992) considers S. spinosus a synonym of S. marisrubri but lists S. zonalis and S. linguafelis as distinct species for the Red Sea. Since the taxonomy and nomenclature of Red Sea spondylids remain unresolved, this article follows the classification of Dekker and Orlin (2000) and two Spondylus spp. are recognised for Farasan (S. groschi and S. spinosus).
Notwithstanding the above, this study produced no new bivalve records for the Red Sea. This is not surprising, given that the marine mollusc fauna of the Red Sea is among the best studied in the world. The checklist of Dekker and Orlin (2000) includes 420 bivalve species for the Red Sea, whereas Dekker and de Ceuninck van Capelle (1994) list 231 bivalve species for the Red Sea coast of Yemen. The OBIS Indo-Pacific Molluscan Database (Rosenberg et al., 2004) reports only 62 bivalve species from the Red Sea and 27 from Saudi Arabian waters. The compilation of Saudi Arabian marine molluscs provided by Sharabati (1981) quotes about 100 mollusc species for the Red Sea and the Arabian Gulf together. With 117, this survey has increased the number of bivalve species recorded for Saudi Arabia by more than three-fold.
As might be expected from a coastline surrounded by tropical coral reefs, the number of bivalves found along beaches with fringing reefs (84 species) was considerably higher than on beaches with fringing mangroves (7 species. In the other two habitats considered, i.e. the pearl oyster community (Ras Shida) and the macro-algal beds and reefs (Qummah and As Saqid West), 46 and 25 bivalve species were found, respectively. These data confirm the species richness of bivalves on coral reefs but also emphasise the species richness of bivalves on secondary hard substrates such as oyster and mussel beds (Mohammed, 1976; Dharmaraj and Chellam, 1983). Relatively high species richness was found at Ras Abrah, Abrah and Al Gora beach, and Ras Farasan (Figure 1). These locations neighbour fringing coral reefs and are relatively exposed to winter winds and currents which deposit large numbers of shells on south-facing beaches. Bivalve species richness was similarly high on Qummah Island (macro-algal beds and reefs) and at Ras Shida, where pearl divers have deposited the remains of biofouling molluscs growing on pearl oyster beds.
Species richness is usually referred to as the number of species present in an ecosystem or a certain location. In practice, measuring the total species richness in an ecosystem is considered impossible, except in very depauperate systems. The observed number of species is a biased estimation of the true species richness in a system, and the observed species number increases non-linearly with sampling effort (Shultz et al., 1999). The species richness as presented in this study should therefore be viewed with caution and treated as a tentative measure of observed species richness in order to compare different locations and habitats on the Farasan Islands. Nevertheless, the comparatively rich bivalve fauna of the Red Sea coast in general, and the Saudi Arabian coast in particular, justifies more detailed studies on the distribution and ecology of bivalves in this region.
Acknowledgements
My gratitude extends to H.H. Prince Bandar bin Saud bin Mohammed Al Saud, Secretary General, Saudi Wildlife Commission, Saudi Arabia, for his continued support towards conservation efforts in Saudi Arabia and for permission to carry out biodiversity research on the Farasan Islands. My appreciation goes also to Dr Tom Butynski (Director, KKWRC) for commenting on an earlier draft of this article, as well as to Ali Sedan (Ranger, Farasan Marine-Protected Area) for his dedication to collecting shells.
Much of the material collected as part of this work has been presented to the Zoological Museum in Hamburg, Germany.
References
Bemert, G. and Ormond, R. (1981) Red Sea Coral Reefs. Kegan Paul International, London, Boston.
Bosch, D.T., Dance, S.P., Moolenbeek, R.G. and Oliver, P.G. (1995) Seashells of Eastern Arabia. Motivate Publishing, Dubai, Abu Dhabi, London.
Dekker, H. and de Ceuninck van Capelle, F.G. (1994) Survey of Yemen Red Sea shells collected by the Tibia-I expedition, 1993. De Kreukel 30 (7–10): 79–147.
Dekker, H. and Goud, J. (1994) Review of the living Indo-West-Pacific species of Divaricella sensu auct. With descriptions of two new species and a summary of the species from other regions. Vita Marina 42: 115–136.
Dekker, H. and Orlin, Z. (2000) Check-list of Red Sea Mollusca. Spirula 47 (supplement): 1–46.
Delsaerdt, A. (1986) Red Sea malacology. Revision of the Chamidae of the Red Sea. Gloria Maris 25: 73–125.
Dharmaraj, S. and Chellam, A. (1983) Settlement and growth of barnacle and associated fouling organisms in pearl culture farm in the Gulf of Mannar. Symposium on Coastal Aquaculture, 12–18 Jan. 1980. Part 2. Molluscan culture. Symposium Serials of the Marine Biology Association of India 6: 608–613.
Gladstone, W. (1994) The Farasan Marine Protected Area. Vol. 1: Biological resources, conservation values, human uses and impacts. NCWCD, Riyadh.
Gladstone, W. (2000) Farasan Islands Protected Area master management plan. NCWCD, Riyadh
Gladstone, W. (2000a) The ecological and social basis for management of a Red Sea marine-protected area. Ocean & Coastal Management 43: 1015–1032.
McFayden, W.A. (1930) The geology of Farasan Islands, Gizan and Kamaran Island, Red Sea. Geological Magazine: 310–332.
Mohammed, M.B.M. (1976) Relationship between biofouling and growth of the pearl oyster Pinctada fucata (Gould) in Kuwait, Arabian Gulf. Hydrobiologia 51: 129–138.
Oliver, P.G. (1992) Bivalved Seashells of the Red Sea. Crista Hemmen, National Museum of Wales, UK.
Oliver, P.G. and Chesney, H.C.G. (1997) Taxonomy and descriptions of bivalves (Lucinoidea, Galleommatoidea, Carditoidea, Cardioidea, Tellinoidea and Myoidea) from the Arabian Sea. Journal of Conchology 36: 51–76.
Oliver, P.G. and Zuschin, M. (2000) Additions to the bivalve fauna of the Red Sea with descriptions of new species of Limopsidae, Tellinidae and Semelidae. Journal of Conchology 37: 17–37.
Rosenberg, G., Ponder, W.F., Middelfart, P.U., Gosliner, T.M., Bouchet, P. and Morris, P.J. (2004) OBIS Indo-Pacific Molluscan Database. URL <http://data.acnatsci.org/obis>.
Rusmore-Villaume, M.L. (2008) Seashells of the Egyptian Red Sea. American University in Kairo Press, Kairo, New York.
Sharabati, D. (1981) Saudi Arabian sea shells. VNU Books International, London.
Sharabati, D. (1984) Red Sea shells. KPI Ltd, London.
Schultz, S.M., Dunham, A.E., Root, K.V., Soucy, S.L., Carroll, S.D. and Ginzburg, L.R. (1999) Conservation Biology. Sinauer Associates, Sunderland, Massachusetts, USA.
Vidal, J. (1997) Large Trachycardiinae from the Indo-West Pacific: The group of Vasticardium orbita (Broderip & Sowerby, 1833) (Mollusca, Cardiidae). Molluscan Research 18: 11–32.