When identifying shells, conchologists often rely, perhaps to an even greater extent than they consciously realise, on colour and colour patterning. But colour and colour pattern are often lost in fossil shells and shells from archaeological sites. In a recent paper (Dommergues E., Dommergues J.- L. and Dommergues C.H. 2006), three of the authors of this note report on the results of a study in which they asked what result the loss of colour information might have on the study of fossil assemblages by applying palaeontological approaches to samples of modern shells without using colour information.
They chose two species of cowry which will be well-known to most readers of Mollusc World: Trivia arctica (Pulteney, 1799) and Trivia monacha (da Costa, 1778). These are clearly distinct species. Fresh shells are clearly and easily separated using colour: T. monacha has three brown dorsal spots; T. arctica has no spots (Fig.1).They also show clear differences in characteristics of the radula and mantle colouration, and T. monacha breeds during late spring and summer and T. arctica in the autumn, winter and early spring (Lebour 1933). Their distributions overlap broadly; populations of T. arctica considered in the study ranged from western Norway to Sicily, and populations of T. monacha from Scotland to Mediterranean France (Fig. 2). Where they are sympatric, they live in similar environments, and individuals of the two species have been found within centimetres of each other in the Atlantic.

Figure 1: Trivia arctica and Trivia monacha. Their shells are very similar apart from the dorsal spots in T. monacha. |
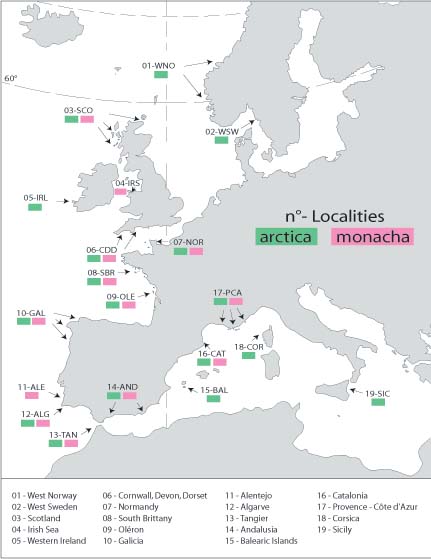
Figure 2: Localities for shells used in the study. |
The first question addressed by the authors of the study was whether they would have realised that two species were present if they studied mixed samples without using the identifications provided by the spots. To do this, they took samples from localities across the range of the two species, some of both species mixed, some of one species only, and looked at three characteristics:
These are basically standard palaeontological methods, though the shape analysis uses a novel approach. The results (Fig. 3) give little indication that two species are present: taking all the shells of both species (almost 1000), the distributions of the values of axis 1 and axis 2 are essentially statistically normal, with no sign of any separation or even of significant bimodality; and the distribution of rib density values is skewed, but there is no indication of significant bimodality.
|
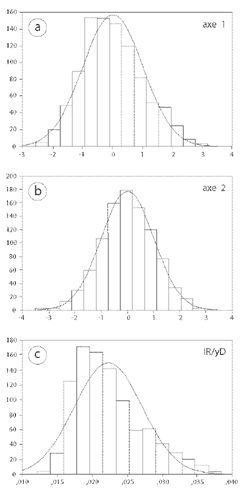
Figure 3: Combined data for both species from all localities show no indication of significant bimodality in two shape indices (a and b); the distribution for rib density ((c): IR/yD) is again not bimodal, but is clearly skewed.
|
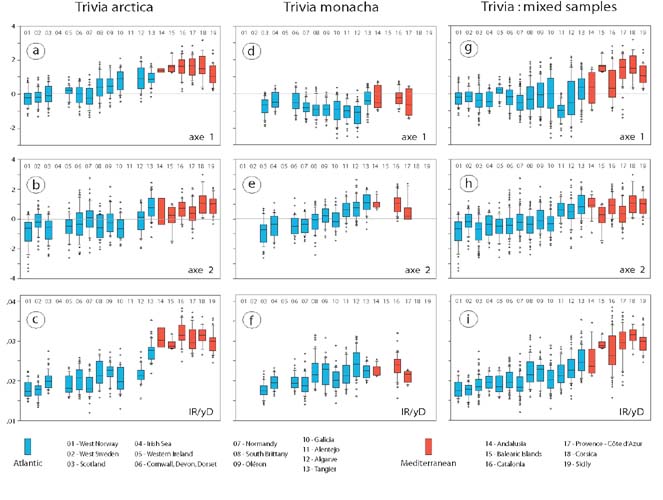
Figure 4: There are gradual (clinal) changes from the N. Atlantic (localities 01-02) to the Mediterranean (localities 14- 19) in T. arctica and T. monacha, and in combined samples. The fairly clear step increase in rib density in T. arctica from the Atlantic to the Mediterranean is less clear in the combined samples.
|
A second part of the study (Fig. 4) looked at variation in different parts of the geographical distribution. When the two species are considered separately (Fig. 4 a-f), both show broadly similar trends,
- in T. arctica (Fig. 4 a-c) values of axes 1 and 2 increase southward, and rib density shows a fairly sharp increase between the Atlantic and the Mediterranean populations (a sample from Tangiers is intermediate);
- in T. monacha (Fig. 4 d-f) values of axis 1 show no very clear trend, values of axis 2 increase southward, and rib density shows a slight increase southward but with no suggestion of a break between Atlantic and Mediterranean populations.
If the species cannot be separated, and so have to be combined for each locality, the results show similar trends but with greater irregularity, and, as Fig. 4 g-i shows, the step in rib density shown by T. arctica between the Atlantic and the Mediterranean is not evident from the combined dataset.
Mixed samples can sometimes be detected because they have greater variability than single-species samples. As Fig. 5 shows, some of the mixed samples show substantially greater variability in the standard deviation of axis 1 values and in the coefficient of variation of rib density, because mean values for the two species are different (though not sufficiently to produce any bimodality). However, in other mixed samples variability is not increased because mean values for the two species are more similar.
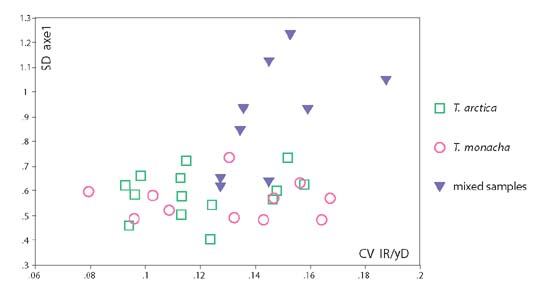 Figure 5:
Figure 5:Mixed samples can sometimes be detected by increased variability, especially in SD axe 1.
This work shows that working without colour information increases the risk of failing to recognise and separate different species, and can create a misleading picture of geographical variation, caused in this case partly by the effects of averaging in mixed samples, and partly by the relative scarcity of T. monacha in the Mediterranean samples. Increased variability can sometimes be used as evidence that a sample contains more than one species; however this requires detailed and sometimes complicated measurements, and does not always detect that a sample includes more than one species.
Acknowledgements:
Many thanks to Christine Street for constructive comment; and to Tim Brooks for help with Fig. 1.
References
Dommergues E., Dommergues J.-L. and Dommergues C.H. (2006), Deux espèces sous un même masque. Le point de vue paléontologique piégé par les coquilles de deux espèces européenes de Trivia (Mollusca, Gastropoda). Revue de Paléobiologie, Genève, 25 (2): 775-790.
Dommergues E., Dommergues J.-L , Magniez F., Neige P. and Verrecchia E.P. (2003), Geometric measurement analysis versus Fourier series analysis for shape characterisation using the gastropod shell (Trivia) as an example. Mathematical Geology 35: 887-894.
